
Grenzen und Ordnung
Ein Schulhof in der großen Pause: Kinder und Jugendliche scheinen ungeordnet durcheinander zu laufen. Einige bilden Gruppen, die stehen oder sich langsam bewegen, andere rennen kreuz und quer, wieder andere gehen gemächlich einzeln oder zu zweit, sitzen auf Banketten oder auf dem Boden. Schaut man dem Treiben länger zu, erkennt man Regeln und Strukturen. Es gibt die sich lebhaft unterhaltenden Gruppen älterer Schüler, ebenso Mädchenzirkel oder auch einen handgreiflichen Streit mit Zuschauern, Pärchen und Einzelgänger, jüngere und ältere Schüler und Schülerrinnen, die sich räumlich streng getrennt aufhalten. Man kann feststellen, dass es eine unsichtbare, aber allen bekannte Kompartimentierung des Schulhofes gibt, die dem bunten Treiben deutliche Grenzen setzt. Dabei kann man zwischen räumlichen (Klassenraum, bestimmter Bereich des Pausenhofes, Lehrerzimmer…) und nicht räumlichen Kompartimenten (Jahrgänge, Klicken) unterscheiden.
Betrachtet man das Innere einer Zelle mit einem starken Lichtmikroskop, kann man auch scheinbar Ungeordnetes beobachten, nicht zuletzt die zitternde Bewegung kleiner Cytoplasmabestandteile, die Robert Brown 1827 als „Molekularbewegung“ erklärt hat. Die exakte Beobachtung durch immer besser auflösende bildgebende Verfahren, Färbungen und Markierungen hat jedoch gezeigt, dass die „Protoplasten“ ein in viele definierte Kompartimente aufgeteiltes sehr komplexes System darstellen.
Immer wenn Wechselwirkungen und Austauschvorgänge eingeschränkt werden, entstehen abgegrenzte Bereiche, in denen mehr solche Austauschvorgänge stattfinden, als in den Bereichen außerhalb des Kompartiments. Die Einschränkungen können auf verschiedene Weise stattfinden. Neben physischen Barrieren können dies auch Bindungen sein, die freie Beweglichkeit verhindern: Man kann einen Hund am Streunen hindern, indem man ihn in einen Käfig sperrt, aber auch indem man ihn an eine Kette legt. Schließlich ist auch in der Biologie eine Kompartimentierung nicht nur räumlich zu denken. Auf molekularer Ebene gibt es bestimmte Markierungen von Molekülen oder Zellorganellen, die Kontakte und Wechselwirkungen begrenzen, z.B. bei der selektiven Wirkung von Hormonen oder Neurotransmittern. Andere nichträumliche Kompartimente sind biologische Arten und die Unterscheidung zwischen „eigen“ und „fremd“.
Im Zustand maximaler Unordnung oder maximaler Entropie gibt es keine Barrieren. Demgegenüber bedeutet Kompartimentierung Ordnung, aber auch unterschiedlich intensive Wechselwirkungen, Hierarchien, Netze, Transportsysteme, Informationen, Informationsverarbeitung, Steuerung und Regelung. Solche Ordnung durch Abgrenzung ist charakteristisch für den gesamten Kosmos. Hier soll es jedoch um die Kompartimente des Lebens und der Lebewesen gehen. Leben wie wir es kennen, ist an einzelne Lebewesen, an Individuen gebunden, die von ihrer Umwelt deutlich abgegrenzt sind. Individuen sind die „Grundkompartimente“ des Lebendigen. Aber jeder Organismus ist auch in seinem Inneren in mehr oder weniger abgeschlossene Reaktionsräume unterteilt. Ebenso geht die Kompartimentierung auf Ebenen oberhalb des Individuums weiter.
In der Regel geht man von einer Komplexitätszunahme dieser Grundkompartimente im Laufe der Evolution aus. Prokaryoten sind weniger kompartimentiert als Eukaryoten, Einzeller weniger als Vielzeller usw. Gleichzeitig bedeutet diese Komplexitätszunahme immer auch eine Überschreitung von vorher bestehenden Grenzen: Eukaryoten sind ein Symbioseprodukt verschiedener Prokaryoten, Vielzeller sind Aggregationen von Zellen, zwischen denen mehr Stoff- und Informationsaustausch stattfindet, als zwischen Einzellern. Seltener ist auch eine Reduktion der Kompartimentierung im Laufe der Evolution möglich, z.B. bei manchen Parasiten.
„Vernetzung“ ist nur möglich, wo es abgegrenzte Bereiche gibt, aber eben auch nur, wo diese Grenzen ausreichend offen sind. In Science-Fiction Entwürfen werden immer wieder Visionen entwickelt, in denen durch neuartige und umfassende Vernetzungen „Superintelligenzen“ entstehen (vgl. z. B. Stanislaw Lems „Solaris“, Crightons „Die Beute“ oder Schätzings „Der Schwarm“. Aber auch in der realen Biologie gibt es Beispiele für „extended organisms“ wie Polypenstöcke, Termiten- und Ameisenkolonien und natürlich alle Formen von Symbiosen.
Lebensentstehung
Schon die Entstehung des Lebens aus unbelebten Vorstufen ist mit zusätzlicher Kompartimentierung verknüpft. Die meisten Vorstellungen von der Biogenese gehen davon aus, dass diese Abgrenzung bereits durch Membranen stattgefunden hat, deren Grundaufbau den heutigen Biomembranen ähnelte. Andere Vorstellungen nehmen an, dass die Grenzbereiche selbst Ausgangspunkt der Lebensentstehung waren, z. B. dass sich Lebensmoleküle an mineralische Oberflächen festgeheftet haben und dadurch ein geordneter Ablauf von Stoffwechselreaktionen möglich wurde (vgl. z. B. Wächtershäuser 2000).
Das „Genkonzept“ von der Entwicklung des Lebendigen sieht in den Nucleinsäuren die „Startmoleküle“ des Lebens. Am Anfang standen RNA-Moleküle, die auch als Enzyme wirken können. Aus der Kooperation solcher Nucleinsäuren mit einer zweiten Molekülklasse, den Proteinen, soll sich dann LUCA (Last Universal Common Ancestor), der letzte gemeinsamen Vorfahr aller Lebewesen, entwickelt haben. Eine Suche nach den Genresten von Luca war jedoch bisher nicht sehr erfolgreich. Deshalb wurde die Vorstellung entwickelt, dass es eine Lebensgemeinschaft von Urlebewesen gab, die zwar gegeneinander abgegrenzt waren, und dadurch jeweils einen eigenen Stoffwechsel hatten, aber ihre Gene teilten. Der bis heute weitverbreitete horizontale Genaustausch bei Prokaryoten wäre dann Rest dieses Urzustandes, eines Zustandes, indem es das Kompartiment „Art“ noch nicht gab (Whitfield 2004).
Die inneren Kompartimente der Eucyten
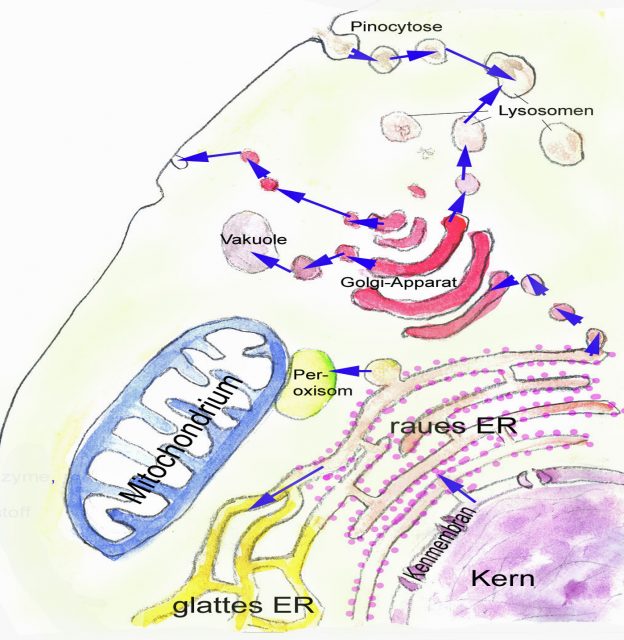
Charakteristisch für die Zellen der Eukaryoten ist, dass sie stark differenzierte innere Membransysteme ausgebildet haben. Die meisten dieser Membransysteme sind entweder unmittelbar miteinander verbunden oder sie stehen über den Austausch von Vesikeln miteinander in Verbindung, Membran umschlossenen Blasen, die sich von Membranen abschnüren oder sich mit Membranen vereinigen können . Zu diesen Membransystemen gehören
- Kernhülle und Endoplasmatisches Retikulum (ER)
- Golgiapparat (Dictyosomen)
- Lysosomen
- Vakuolen
- Peroxisomen bzw. Microbodies
- Plasmamembran (als Abschluss des Zellkörpers nach außen)
Nicht mit den übrigen Membransystemen in Verbindung stehende Kompartimente, die durch Doppelmembranen vom Zytoplasma abgegrenzt sind:
- Plastiden
- Mitochondrien
Eine Erklärung für die Sonderstellung dieser beiden Zellorganelle ergibt sich aus ihrer stammesgeschichtlichen Entstehung aus Endosymbionten. Während die Innenmembran der Mitochondrien stark aufgefaltet ist, enthalten die Plastiden insbesondere die grünen Chloroplasten in ihrem Inneren ein weiteres Membransystem aus sogenannten Thylakoiden, das durch Abschnürung aus der inneren Plastidenmembran entsteht, aber im Endzustand nicht mehr mit ihr verbunden ist. In als Grana bezeichneten Thylakoidstapeln sind die Pigmente und Enzyme der Photosynthese untergebracht. Plastiden sind über dünne, von beiden Membranen umgebene ?lasmakanale, den sogenannten Stromuli untereinander und auch mit Zellkern und Mitochondrien verbunden (Krupinska et al. 2010).
Das zweite Kompartimentierungssystem der Zellen besteht aus fädigen Proteinstrukturen: Aktinfilamenten, Mikrotubuli und intermediären Filamenten. Alle drei stellen Polymere aus kleineren Proteinmolekülen dar (Abb. ). Sie bilden in der Zelle ein netzartiges Gerüst, das auch an der Zellmembran verankert ist. Aktinfilamente sind, oft in Verbindung mit dem Motorprotein Myosin, für Bewegungen der ganzen Zelle – besonders augenfällig bei Muskelzellen – sowie unterschiedliche Viskositätszustände des Cytoplasmas zuständig. Sie können äußere Gestaltänderungen der Zellen bewirken. Mikrotubuli bewegen Zellorganelle durch das Cytosol und Chromosomen bei der Mitose. Sie sind die bewegenden Strukturen von Cilien und Geiseln. Intermediäre Filamente sind sehr stabile seilartige Fadenstrukturen, die z.B. für die Stabilität des Zellkerns und der Nervenfasern und für die Zerreiß- und Zugfestigkeit von Epithelien verantwortlich sind.
Kompartimente bei Prokaryota
Das innere Membransystem der Eucyten ermöglicht die vielfältigen nebeneinander ablaufenden Reaktionen in der Zelle, in dem es Reaktionsräume, Speicherräume und Entsorgungsräume gegeneinander abgrenzt. In Procyten ist das innere Membransystem im Allgemeinen nicht so stark ausgeprägt, wenngleich auch die Zellen vieler Prokaryoten reichlich innere Membranen enthalten, die aus Einstülpungen der Zellmembran hervorgehen. Bei den Cyanobakterien tragen diese intracytoplasmatischen Membranen (ICM) die Pigmente und Enzyme für die Photosynthese, bei aeroben Bakterien sind die Enzyme für die Zellatmung an inneren Membranen verankert. Dafür, dass auch in Prokaryoten viele Stoffwechselreaktionen parallel stattfinden können, ohne sich gegenseitig zu behindern, sind neben Membranabgrenzungen Proteinstrukturen verantwortlich, die im Cytosol oder an der Zellmembran relativ fest verankert sind und Stoffwechselpartner an sich binden.
Es gibt Hinweise, dass die stärkere Kompartimentierung der Eucyten mit dem steigenden Sauerstoffgehalt der Atmosphäre zusammenhängt. Nach Acquisti et al. (2007) sind sauerstoffreiche Membranproteine bei einer reduzierenden Umgebung weniger stabil als sauerstoffarme. Gerade für Signal übertragende Transmembranproteine sind solche sauerstoffreichen Domänen aber charakteristisch. Mit der Erhöhung des atmosphärischen Sauerstoffgehaltes konnte der Einbau solcher Proteine in Biomembranen zunehmen. Dies betrifft insbesondere die für die Signalübertragung durch Membranen nötigen Proteine mit relativ großen auf der Außenseite der Membran liegenden Rezeptorstrukturen.
Struktur und Funktion von Biomembranen
Schon bevor man Biomembranen im Elektronenmikroskop sichtbar machen konnte, ließen bestimmte chemische und physikalische Eigenschaften darauf schließen, dass Lipide ein wichtiger Bestandteil dieser Membranen seien. So beobachtete man, dass fettlösliche Substanzen von den Zellen viel leichter aufgenommen wurden, als wasserlösliche. Außerdem stellte man fest, dass Zellmembranlipide auf Wasser einmolekulare Schichten bilden, um die Zelle aber in einer Doppelschicht vorliegen müssen (Gorter, Grendel 1925 nach Helmich 2001/2005). Dass Zellmembranen auch für Wasser und anorganische Ionen in gewissem Umfang durchlässig sind, kann man sich nur erklären, wenn man annimmt, dass in die Lipiddoppelschicht auch Proteinmoleküle eingelagert sind. Eine der ersten Vorstellungen vom Aufbau der Membranen ging von einer Lipiddoppelschicht, vorwiegend aus Phospholipiden, aus, auf der Proteine aufgelagert sein sollten. In einer Weiterentwicklung dieses Modells gingen Singer und Nicolson 1972 davon aus, dass die Proteinmoleküle in der Lipidschicht wie Eisberge im Meer schwimmen. Einige Proteinmoleküle durchdringen die Lipidschicht (Tunnelproteine, Kanalproteine), sie können passiven oder aktiven Stofftransport und Signalübertragung durch die Membran vermitteln.
Die Vorstellung einer Membran als Flüssigkeitsfilm mit frei beweglichen Proteinen stimmt jedoch nicht ganz, die Proteine sind in ihrer Beweglichkeit durchaus eingeschränkt, dabei kann z. B. das Cytoskelett an der Membraninnenseite eine Rolle spielen, das Bereiche mit bestimmten Proteinmolekülen „einzäunt“. Gleichzeitig wirken bestimmte Proteine wie „Zaunpfosten“ (Fence-and-Picket-Modell, Suzuki 2005). Diese abgegrenzten Bezirke können aber von bestimmten Proteinen auch übersprungen werden (Abbot 2005). Eine andere Modellvorstellung geht davon aus, dass es in den Membranen floßartige Lipidschollen („lipid rafts“) gibt, die zähflüssiger sind und mit ihren Proteinen in dem Lipidfilm driften. Dabei können einzelne Proteine von diesen Flößen aufgenommen oder abgegeben werden (Simons, Ikonen 1997).
Der Aufbau der Lipiddoppelschicht ist für deren Flüssigkeit von Bedeutung. Ungesättigte Fettsäuren in den lipophilen Schwänzen der Phospholipide haben einen Knick, der die Moleküle am dichten Zusammenrücken hindert, und fördern dadurch die Fluidität. In die Lipiddoppelschichten eingebaute Cholesterinmoleküle vermindern bei mäßigen Temperaturen die Membranflüssigkeit, weil sie die Beweglichkeit der Phospholipide einschränken. Bei niedriger Temperatur stören sie jedoch die regelmäßige, dichte Packung und verhindern dadurch, dass die Membranen „kristallisieren“.
Die wichtigsten Funktionen der Membranproteine sind:
- Transport (passiv, aktiv)
- Enzymaktivität
- Signalübertragung
- Verbindung von Zellen
- Zellerkennung
- Verankerung am Cytoskelett und an der extrazellulären Matrix
Neben Proteinen sind für die Zellerkennung auch Membran gebundene Kohlenhydrate von großer Bedeutung (Glykoproteine und Glykolipide).
Vom Einzeller zum Vielzeller
Auch Vielzeller entstehen normalerweise aus einer Zelle. Nach den Mitosen bleiben die Zellen jedoch verbunden und geben damit einen Teil ihrer Selbständigkeit auf. Während sie zunächst noch weitgehend identisch und damit „totipotent“ sind, differenzieren sie sich im Laufe der weiteren Entwicklung und damit können nur noch bestimmte Zelltypen aus ihnen hervorgehen („multipotent“). Schließlich sind sie überhaupt nicht mehr teilungsfähig. Damit ist der natürliche Tod der Zellen der Vielzeller vorprogrammiert.
Dieser Übergang von Einzellern zu Vielzellern , der vor etwa einer Milliarde Jahre stattfand, konnte nur funktionieren, wenn weitere Probleme gelöst wurden. Bei Einzellern läuft die natürliche Selektion zwischen den einzelnen Zellen ab. Sie sind die Einheiten der Selektion. Bei Vielzellern darf es keine Selektion zwischen den Körperzellen geben. Das kann nur gelingen, wenn es einen Erkennungsmechanismus von „eigen“ und „fremd“ gibt. Ein solches Selbsterkennungssystem kann als der Anfang eines Immunsystems aufgefasst werden.
Dieses Selbsterkennungssystem ist bei „niederen“ Vielzellern noch nicht sehr ausgeprägt. Deshalb funktioniert z.B. das Propfen bei Pflanzen – sogar zwischen Individuen verschiedenen Arten – meist sehr gut. Auch bei koloniebildenden Tieren ist das Erkennungssystem im Allgemeinen so, dass es zwischen den Einzelindividuen einer Kolonie nicht unterscheidet. Relativ gut untersucht sind die Verhältnisse bei dem koloniebildenden Manteltier Botryllus schlosseri. Das genetisch verankerte „Selbsterkennungssystem“ dieser Seescheide erlaubt nur die Fusion von genetisch nahe verwandten Kolonien. Von manchen Forschern wird daraus gefolgert, dass der ursprüngliche Sinn des Immunsystems die Verhinderung solcher Zellinvasionen war, die eine Konkurrenz unterhalb des Individuums bewirken würden. Evolutionsbiologisch gesehen könnte man sagen, das Selbsterkennungssystem sorgt bei Vielzellern dafür, dass der Gesamtorganismus und nicht einzelne Zellen oder Zelllinien die Einheit der Evolution sind.
Diese Sicht könnte auch ein neues Licht auf das Wirkungsgefüge von Krebsbildungen werfen. So weiß man heute, dass spezielle Krebsstammzellen für die Krebsbildung und die Metastasenbildung entscheidend sind (Clarke, Becker 2007). Weissmann (Ainssworth 2006) sieht gewisse Parallelen zwischen Krebszellen und den Gewinner-Stammzellen von Botryllus. Er meint, wenn man die Gene der Botryllus-Übernahme-Zellen entschlüsseln würde, würde man wahrscheinlich Ähnlichkeiten bei den Genen finden, die Krebszellen ihre tödliche Entwicklung ermöglichen. Aus dieser Sichtweise könnte man Krebs als ein Relikt bzw. einen Atavismus aus der Zeit des Übergangs von Einzellern zu Vielzellern ansehen.
Zellen und Gewebe
Die Plasmamembran ist die äußere Grenze einer Zelle, aber die meisten Zellen bilden weitere Strukturen aus, die außerhalb der Plasmamembran liegen. Pflanzenzellen z. B. sind von einer festen Zellwand aus Zellulose umgeben. Bei Pilzen besteht diese Zellwand aus Chitin. Die Zellen vielzelliger Tiere besitzen zwar keine den Pflanzenzellen vergleichbare Zellwände, sie verfügen aber über eine hoch entwickelte extrazelluläre Matrix, die vorwiegend aus von der Zelle abgesonderten Proteinfasern (Kollagene, elastische Fasern) und einer Grundsubstanz aus Glykosaminglykanen, Proteoglykanen und Adhäsionsproteinen (Glykoproteinen) besteht. Diese extrazelluläre Matrix ist mit Proteinen der Zellmembran verbunden (Integrine) und über diese in die Membran integrierten Proteinmoleküle ist auch ein Signalaustausch von der extrazellulären Matrix in das Cytosol der Zelle hinein möglich.
In einem Verband aus vielen Zellen (Gewebe) kann die extrazelluläre Matrix koordinierende Signale übertragen. Dies spielt eine wichtige Rolle bei der embryonalen Gewebe- und Organentwicklung, aber auch bei der Tumorbildung. Dabei spielt die Basallamina als besondere Ausbildung der Extrazellulären Matrix, die Zellen und Epithelien von umgebenden Bindegeweben trennt, eine wichtige Rolle.
In vielzelligen Tieren und Pflanzen sind viele Einzelzellen zu funktionsfähigen Geweben und Organen verbunden. Durch spezielle Plasmaverbindungen können nicht nur kleine Moleküle, Wasser und Ionen sondern auch Proteine und RNA-Moleküle ausgetauscht werden. Für den Transport dieser größeren Moleküle sind Cytosklelettfasern verantwortlich. Bei Pflanzen nennt man diese Verbindungen Plasmodesmen. Bei Tieren gibt es verschiedenen Typen von Zellverbindungen. Besonders häufig sind solche Zellverbindungen in Epithelgeweben, welche die inneren und äußeren Oberflächen eines Tierkörpers auskleiden. Gap Junctions (Kommunikationskontakte) bilden winzige Cytoplasmakanäle zwischen benachbarten Tierzellen. Durch diese Kanäle können Salze, Zucker, Aminosäuren und andere kleine Moleküle bis zu einem Molekulargewicht von 2.000 diffundieren. Weitere Zellverbindungen sind Tight Junctions oder Verschlusskontakte, die Epithelzellen gürtelartig verbinden und verhindern, dass extrazelluläre Flüssigkeit durch ein Epithel hindurchsickert. Im Gehirn bilden die dichten Tight Junctions zwischen den Endothelzellen der Blutkapillaren die Blut-Hirn-Schranke. Desmosomen und Adhärenz-kontakte („Haftkontakte“) wirken nietenartig und verbinden verschiedene Zellen zu einer Gewebeschicht.
Von Geweben zu Organen
Gewebe setzen sich aus einheitlichen Zellen zusammen, verschiedene Gewebe sind im Tierkörper zu Organen zusammengefasst. Die verschiedenen Organe stehen zwar in enger Wechselwirkung miteinander, durch die starke Abgrenzung dieser Einheiten sind aber verschiedene Funktionen wie Verdauung, Atmung, Blutkreislauf oder Exkretion erst möglich. Solche Organe bilden als Ganzes relativ abgeschlossene Systeme im Organismus und erst dadurch wird z. B. die Organtransplantation möglich.
Größere Organismen benötigen zum Stofftransport spezielle Transportsysteme. Bei Pflanzen handelt es sich dabei überwiegend um Durchflusssysteme, bei Tieren um Kreislaufsysteme. Auch diese Systeme müssen vom übrigen Körpergewebe mehr oder weniger abgeschlossen sein, um einen wirkungsvollen Stofftransport zu ermöglichen. Aber auch offene Kreislaufsysteme, wie es z. B. für die große Gruppe der Insekten charakteristisch sind, können sehr effektiv arbeiten.
Ein besonders wichtiges, stark kompartimentiertes Stoffwechselorgan des menschlichen Körpers und des Körpers der Wirbeltiere ist die Leber. Beim Menschen liegt sie im oberen Teil der Bauchhöhle unmittelbar unter dem Zwerchfell Sie ist mit einer Masse von rund 2 Kilogramm die größte Körperdrüse. Pro Minute wird sie von einem Liter Blut durchflossen.
Die diffizile Kompartimentierung der Leber gestattet, dass mehr als 500 verschiedene Stoffwechselvorgänge hier stattfinden können. Zunächst sorgen zwei Zufluss- und zwei Abflusssysteme dafür, dass sich in den Hepatocyten die richtigen Konzentrationsgefälle einstellen können, die für die Umbaureaktionen Voraussetzung sind:
- Die Pfortader stellt die Verbindung zum Darm her und sorgt dafür, dass die vom Darm resorbierten Nährstoffe zur Leber gelangen.
- Über die Leberarterie werden den Hepatocyten Sauerstoff und Signalstoffe, aber auch Aufbaustoffe zugeführt.
- Über die Lebervene werden Abfallstoffe zur Ausscheidung durch die Niere und CO2 zu Abgabe in der Lunge abtransportiert.
- Auch mit Gallenflüssigkeit werden Abfallstoffe über die Gallengänge und die Gallenblase zum Dünndarm abtransportiert., z.B. die Abbauprodukte des Häms, die gelben Bilirubine.
Die Leber besteht aus einem größeren rechten und einem kleineren linken Lappen, die sich jeweils in Tausende Leberläppchen unterteilen. In der Mitte jedes dieser Läppchen von etwa 1 mm Durchmesser liegt eine kleine Zentralvene, die das Blut zur Lebervene leitet. Zwischen den Läppchen liegen Bindegewebsfelder, durch die sich je ein feiner Ast der Leberschlagader und der Pfortader zeiht, deren Blut durch das Leberläppchen zur Sammelvene sickert, sowie ein Gallenkanälchen, das im Läppchen produzierte Gallenflüssigkeit in zum Blutstrom entgegen gesetzter Richtung zur Gallenblase abtransportiert. Das kleinste Kompartiment des Organs Leber ist die Leberzelle (Hepatozyt). Die Leberzellen sind lamellenartig angeordnet und lassen Kanälchen frei, durch die das Blut sickern kann (Sinusoide).
In den Leberzellen werden viele Eiweißmoleküle aufgebaut, außerdem werden Giftstoffe und Stoffe, die aus dem Körper befördert werden, sollen für die Ausscheidung vorbereitet. Die Glucose aus der Verdauung der Kohlenhydrate kann in den polymeren und damit osmotisch unwirksamen Speicherstoff Glykogen umgewandelt werden, der zum Teil in der Leber selbst gespeichert werden kann. Verschiedene Lipide werden in der Leber aus ihren Bestandteilen aufgebaut und umgebaut, u. a. das Cholesterin.
Bei dem Abbau und Umbau von stickstoffhaltigen Proteinen wird mehr Stickstoff frei als für den neuen Eiweißaufbau benötigt wird. Dieser überschüssige Stickstoff wird in der Leber in Harnstoff umgewandelt, ein Sekretionsprodukt, das an die Lebervene abgegeben, von den Nieren herausgefiltert und mit dem Urin ausgeschieden wird. Auch die Gallenflüssigkeit wird in der Leber produziert und durch besondere Gallengänge zur Gallenblase befördert, von der aus sie über den Gallengang in den Dünndarm ausfließt. Sie besteht aus Gallensäuren bzw.-salzen, Lipiden, Cholesterin und Farbstoffen. Diese Farbstoffe sind Abbauprodukte des Hämoglobins, die sogenannten Bilirubine. Sie sind sowohl für die Gelbfärbung der Gallenflüssigkeit und des Urins als auch für die Braunfärbung des Stuhls verantwortlich. Die Gallensäuren sind Abbauprodukte des Cholesterins. Sie helfen als Emulgatoren bei der Fettverdauung im Dünndarm. Wird der Abfluss der Gallenflüssigkeit verhindert – z.B. durch Gallensteine oder eine Leberentzündung – kann der Bilirubinüberschuss im Blut eine Gelbsucht bewirken.
Durch Schädigungen, wie sie z. B. durch reichlichen Alkoholkonsum hervorgerufen werden können, kann es zur sogenannten Leberzirrhose kommen. Dabei handelt es sich um eine teilweise Zerstörung der Leberzellen. Abgestorbene Hepatozyten werden durch Bindegewebe ersetzt, welches das Organ durchzieht und noch funktionsfähige Zellen isoliert. Diese isolierten Inseln sind von den Zufuhr- und Abfuhrsystemen mehr oder weniger abgetrennt und können deshalb die vielseitigen Stoffwechselaufgaben nicht mehr erfüllen. Dadurch, dass weniger Pfortaderblut aufgenommen werden kann, kommt zu einem Rückstau. Durch die Gefäßwände wird Wasser in die Leibeshöhle filtriert. Der Abtransport der Gallenfarbstoffe über die Gallenblase wird behindert, wodurch es zu gelbsuchtartigen Zuständen kommt usw.
Kompartimente oberhalb des Individuums
Auch oberhalb der Organisationsebenen Zelle, Gewebe, Organ, Organismus ist die Biosphäre in viele Kompartimente gegliedert, wie Populationen, Arten, Biozönosen, Ökosysteme, Biome, Reviere, Areale usw. Dabei wird hier noch deutlicher, dass es neben räumlich definierbaren Kompartimenten auch Kompartimente gibt, die sich aus den besonderen Eigenschaften ihrer Bestandteile ergeben: Populationen und Arten sind durch den gemeinsamen Genpool und die Fähigkeit zum Genaustausch (Sexualität) gekennzeichnet. Räumlich kann sich eine Population aber über den ganzen Erdball erstrecken. Reviere und Areale können sich räumlich vielfach überlappen und überdecken. Derselbe geographische Raum kann z.B. viele Reviere verschiedener Arten enthalten. Ein geographischer Raum mit vielen gemeinsamen Artarealen wird als Floren- oder Faunenregion bezeichnet.
Besonders einschneidend ist die Grenze, die einen Organismus bzw. ein Individuum gegen seine Umwelt abgrenzt. Kreislaufsysteme überschreiten diese Außengrenze ebenso wenig, wie Zellen mit dem speziellen genetischen Programm, das nur für dieses Individuum gilt. Ein spezielles Signalsystem sorgt für die Koordination aller Zellen, Gewebe und Organe innerhalb des Individuums, nur wenig davon dringt nach außen. Innerhalb des Organismus wird durch aufeinander abgestimmte Stoffwechselvorgänge ein stoffliches Gleichgewicht aufrecht erhalten, das man mit einem eigenen Begriff „Homöostase“ kennzeichnet und das die Grenzen des Organismus nicht überschreitet. Wenn die genannten individuellen Schranken überschritten werden, nehmen wir das als etwas Besonderes wahr: Bei Säugetieren ist der Kreislauf des Muttertieres mit dem Embryo verbunden, bei Kolonie bildenden oder Staaten bildenden Tieren sind die Individualgrenzen ebenfalls mehr oder weniger stark aufgelöst.
Auch der Sexualvorgang ist eine besondere Grenzüberschreitung, durch die gleichzeitig ein höheres Kompartiment gebildet wird, die Gemeinschaft aller Individuen, zwischen denen Gene ausgetauscht werden können, die Art. Die individuellen genetischen Programme machen die innerartliche Evolution möglich, die Genpools der Populationen und Arten sind die Grundlage für die Evolution oberhalb des Artniveaus.
Die Individuen, die zu einer Art gehören, haben in der Regel ähnliche Ansprüche an ihre Umwelt. Im Bezug auf bestimmte Umweltfaktoren spricht man vom „Toleranzbereich“ der Art. Diese verschiedenen Toleranzbereiche beschränken die Verbreitung der Art. Die räumliche Verbreitung, das Artareal, wird aber auch durch erdgeschichtliche Entwicklungen bestimmt. Dazu gehören tektonischen Vorgänge, insbesondere Verschiebungen der Kontinentalplatten, Gebirgsbildungen und Überflutungen (Meerestransgressionen), Klimaeinbrüche und in der Folge auch Konkurrenzbeziehungen zu Arten, zu denen vorher kein Kontakt bestand.
Arten stehen in vielen Wechselbeziehungen mit der Umwelt und mit anderen Arten. Vorwiegend durch geografische und geologische Gegebenheiten werden diese Wechselbeziehungen aber beschränkt und gelenkt. In bestimmten Gebieten und zwischen den darin vorkommenden Arten sind die Wechselbeziehungen vielfältiger als nach außen. Die Folge ist, dass sich die Biosphäre abgestuft in viele Teilräume untergliedern lässt, die allgemein als Ökosysteme bezeichnet werden. Ein solches Ökosystem kann ein kleines Feldgehölz, eine Weidetümpel oder eine Blockhalde an einem Bergsturz sein, aber auch der Amazonas-Regenwald, das Kongobecken, die circumpolare Tundra oder ein Ozean. Großökosysteme, die sich in viele Teilsysteme untergliedern, werden auch Biome genannt (Whittaker 1975, Walter 1976, UB 299). Sind sie vorwiegend von den Klimazonen der Erde bestimmt, nennt man sie Zonobiome, in den verschiedenen Höhenstufen der Gebirge unterscheidet man Orobiome, besondere Bodenbedingungen führen zu speziellen Pedobiomen.
Kleine Ökosysteme, die eine Landschaft untergliedern, werden oft auch als „Biotope“ bezeichnet, obwohl dieser Begriff in der ökologischen Terminologie ursprünglich nur den Lebensraum ohne die Lebensgemeinschaft bezeichnet. Der Begriffswandel lässt sich aus der Naturschutzpraxis erklären: Wenn man ein bestimmtes Ökosystem durch Naturschutzmaßnahmen einrichten will, muss man zunächst die standörtlichen Bedingungen schaffen. So „legt man einen Biotop an“ – z.B. einen Gartenteich oder eine Natursteinmauer –, der dann durch Bepflanzung oder natürliche Ansiedlung von Arten zum Kleinökosystem wird. Oft wird mit dem Begriff „Biotop“ auch gleich ein bestimmter ökologischer Wert verbunden. „Biotopkartierungen“ in der Kulturlandschaft erfassen in der Regel nur besondere, „ökologisch wertvolle“, „naturnahe“ Landschaftselemente.
Eine andere Kompartimentierung der Landschaft ergibt sich aus den Revieren verschiedener Tierarten. Die Grenzen werden hier vorwiegend durch das agonistische Verhalten der Revierbesitzer errichtet. Aber auch spezielle akustische, optische oder chemische Signale wirken begrenzend.
Grenzen in Naturlandschaften sind oft nicht sehr scharf, sondern durch Übergänge gekennzeichnet, die man mit einem eigenen Begriff erfasst: Ökotone. So ist es oft nicht möglich, die Grenzen zwischen zwei Ökosystemen (oder zwei Pflanzengemeinschaften) genau festzulegen. In Kulturlandschaften sind die Grenzen jedoch in der Regel scharf, da sie durch menschliche Aktivitäten bedingt sind. Sehr gut lässt sich dies von Flugzeug aus oder an den Bildern von Google Earth erkennen. So ist es auch kein Wunder, dass die durch die „Pflanzenoziologie“ gekennzeichneten Pflanzengesellschaften vor allem für Mitteleuropa zu einem sehr differenzierten System ausgebaut wurden. Allerdings wird sich „ein mehr Außenstehender … die Frage aufwerfen, ob die Katalogisierung aller, auch der kleinsten Vegetationseinheiten Mitteleuropas die dafür aufgewendete Mühe lohnt. Dies wäre vom wissenschaftlichen Standpunkt aus durchaus zu bejahen, wenn die derzeitigen Pflanzengesellschaften ähnlich unveränderliche Einheiten wären wie die taxonomischen, aber das sind sie nicht“ (Walter 1973, S.115).
In den heutigen Kulturlandschaften ist die „Überkompartimentierung“ ebenso ein Naturschutzproblem wie die „Unterkompartimentierung“ durch riesige Monokulturen. Von einem durch Ackerflächen umschlossenen Kleinkompartiment „Feldgehölz“ aus ist es z. B. für viele Tiere schwierig, in andere, ähnliche Biotope zu gelangen. Feldhecken begrenzen Kulturflächen, sie sind aber auch Verbindungswege zwischen Ökosystemen. Besonders stark wirkende Grenzen sind Verkehrswege, weshalb man an einigen wenigen Stellen sinnvoller Weise so genannte Biotopbrücken über Autobahnen gebaut hat, um deren Landschaft zerschneidende Wirkung zu mindern.
Das „Basiskonzept „Kompartimentierung“ im Unterricht
„Lebende Systeme zeigen abgegrenzte Reaktionsräume. Dieses Basiskonzept hilft z. B. beim Verständnis der Zellorganellen, der Organe und der Biosphäre“. So steht es in der „Einheitlichen Prüfungsanforderung in der Abiturprüfung Biologie“ nach dem Beschluss der KMK-Konferenz vom 05.02.2004. In Lehrbüchern tritt der Begriff jedoch meistens nur im Zusammenhang mit der „Zellkompartimentierung“ auf, seltener auch im Zusammenhang mit der „Kompartimentierung des Organismus“ (z.B. Biesalski, Grimm 2002).
Damit wird die Intention der „Basiskonzepte“ oder „Erschließungsfelder“ nicht erfüllt. Denn dadurch, dass Basiskonzepte biologische Phänomene umreißen, die in der Regel durch viele, wenn nicht alle Organisationsebenen des Lebendigen hindurchreichen, sollen sie biologische Fachkenntnisse strukturieren und dadurch fassbarer und merkbarer machen.
Wie könnte gerade das Basiskonzept „Kompartimentierung“ helfen, Lernen zu verbessern? Wie könnte es kumulatives und outputorientiertes Lernen fördern?
Die wichtigste Gemeinsamkeit der Kompartimente auf allen biologischen Organisationsebenen ist die selektive Abgrenzung. Dies betrifft den Austausch von Stoffen, von Energie und von Information. Diese Einschränkungen können aber – ähnlich wie eine Zollstation und eine Grenzkontrolle an einer Ländergrenze – zur Steuerung und Regelung, auch zur gezielten Signalweitergabe genutzt werden.
Damit hat man ein strukturierendes Prinzip für viele biologische Sachverhalte gewonnen, das so unterschiedliche Inhalte, wie „Stoffkreisläufe in Ökosystemen“ und „Intrazelluläre Regelprozesse“ oder „Biomembranen“ und „Vernetzung von Biotopen“ in Beziehung bringen kann. Gleichzeitig kann man neue Inhalte mit diesem Prinzip aufschließen, erklären, besser verstehen und einordnen (Outputorientierung).
| Kompartiment | Grenze für | Abgrenzung durch | Grenzüberwindung durch |
| Membranumschlossenes Zellkompartiment | Moleküle, Ionen | Lipiddoppelschicht | Tunnel- und Carrierproteine, signalübertragende Proteine, lipophile Moleküle |
| Organ | Blut, Lymphe u.a. Körperflüssigkeiten | Epithelien | Blutgefäße, Lymphe |
| Organismus | Stoffe, Energie, Signale | Haut, Epithelien | Verdauungssystem, Sinnesorgane, Kommunikationssysteme |
| Art, Population | Gene | Kreuzungsbarrieren, Inkompatibilitätsfaktoren | Migration, Hybridisierung, horizontaler Gentransfer |
| Areal (Verbreitungsgebiet) | Individuen einer Art (oder einer höheren Verwandtschaftsgruppe) | Geographische und geologische Barrieren, Konkurrenzdruck anderer Arten | Verschleppung von Individuen durch natürliche oder vom Menschen verursachte Vorgänge |
| Revier | Individuen | Agonistisches Verhalten; akustische, optische, chemische Signale | Revierkämpfe |
| Ökosystem | Individuen, Stoffe, Energie | Geografische Barrieren | Tierwanderungen, Transport von Vermehrungseinheiten, Stofftransport über Gewässer, Wettergeschehen wie Luftströmungen |
| Biom | Ökosysteme, Arten/Populationen, Individuen | Klimagrenzen, geographische Barrieren | Klimaänderungen, Erosion, Tektonik |
Wenn man erkannt hat, dass Grenzen auch etwas mit Austausch zu tun haben, versteht man das in lebenden Systemen immer wiederkehrende Prinzip der Oberflächenvergrößerung zur Förderung von Austauschprozessen besser. Auch der modulartige Aufbau von Lebensstrukturen kann mit dem Prinzip der Kompartimentierung in Verbindung gebracht werden (Grundorgane der Pflanze, die sich immer wiederholen; Metamerie bei Tieren). So kann dieses Basiskonzept, wie auch andere, helfen, über Querverbindungen vernetztes Lernen zu erleichtern und doch bei dieser Vernetzung Chaos zu vermeiden. „Alles hängt mit allem zusammen“, ist zwar eine korrekte Beschreibung der Welt, verhilft aber kaum zu einem besseren Weltverständnis.
In diesem Artikel sind wir von den kleinsten Kompartimenten des Lebendigen in den Zellen ausgegangen und haben uns dann über Organe und Organismen zu den Überindividuellen Kompartimenten der Biosphäre emporgearbeitet. Dies muss aber nicht der Weg sein, der sich auch für den Schulunterricht anbietet. Die frühe Behandlung cytologischer und sogar molekularbiologischer Inhalte führt zwangsläufig dazu, dass die „organismische Biologie“ an Bedeutung verliert. Gerade in den Klassenstufen 5 bis 7 hat die unmittelbare Begegnung mit Tier- und Pflanzenarten, möglichst in ihren natürlichen Lebensräumen, einen besonders animierenden und prägenden Einfluss. Dies ist nämlich der Zeitabschnitt, in dem sich bei mangelnder Förderung das Interesse an der „grünen Biologie“ allmählich verliert. Ziel einer ausgewogenen Allgemeinbildung sollte es aber sein, Interesse und Kenntnis der „Vielfalt des Lebendigen“ in den makroskopischen Dimensionen zu erhalten und zu fördern. Deshalb wäre es durchaus sinnvoll, von Individuen und Arten ausgehend in den unteren Klassen der SI einen deutlichen Schwerpunkt auf Lebensräume und Ökosysteme zu legen und diese „Landschaftsbiologie“ auch mit Unterrichtsabschnitten im Gelände zu vermitteln. Das würde z. B. bedeuten, dass man wichtige heimatliche Lebensräume wie Fließgewässer und Teich, Hecke und Wald, Wiese und Weide aus eigener Anschauung kennen lernt und dass man möglichkeiten der landschaftsgestaltung im eigenen Schulgarten erfährt.. Cytologische und molekularbiologische Inhalte sollten schwerpunktmäßig auf die letzten Klassenstufen verlegt werden. Die in der Makrobiologie gewonnen Vorstellungen zur Kompartimentierung könnten dann als Modelle für mikroskopische und submikroskopische Vorstellungen dienen. Begriffe wie „Tunnelprotein“, „Fence-and-Picket-Modell“ oder „aktiver und passiver Transport“ bauen ja ohnehin makroskopischen Vorstellungen auf.
Literatur und URLs
Abbot, A.: Cell biology: Hopping fences. Nature 433, p.680-683, 2005
Acquisti, C., Kleffe, J., Collins, S.: Oxygen content of transmembrane proteins over macroevolutionary time scales. Nature 440, p.47-52, 2007
Ainsworth, C.: Cell biology: The Story of I. Nature 440, p. 730-733, 2006
Alberts, B., Bray, D. Lewis, J., Raff, M., Roberts, K., Watson, J.D.: Lehrbuch der molekularen Zellbiologie. Wiley-VCH, 2001
Archibald, J.: One plus one equals one- symbiosis and the evolution of complex life. Oxford University Press 2014
Biesalski, H.K., Grimm, P.: Taschenatlas der Ernährung. Thieme, Stuttgart, 2.A., 2002
Brenner, K.-U.: Der Körper des Menschen. Weltbild, Augsburg 1990
Campbell, N.A., Reece, J.B. (Hrsg. d. dtsch. Ausg. H. Markl): Biologie. Spektrum, Heidelberg/Berlin, 6. A. 2003
Clarke, M.F., Becker, M.W.: Krebs – sind Stammzellen schuld? Spektrum der Wissenschaft , S. 56-63, Januar 2007
Einheitlichen Prüfungsanforderung in der Abiturprüfung Biologie. Beschluss der Kultusministerkonferenz vom 01.12.1989 i. d. F. vom 05.02. 2004
Faller, A., Schünke, M.: Der Körper des Menschen. Thieme, Stuttgart 1999
Frey, W., Lösch, R.: Lehrbuch der Geobotanik. Elsevier, München 2.A. 2004
Graf, D.: Nano-Katastrophen (Michael Crighton: Die Beute). UB Kompakt 312 (Jg. 30), S.25-28, 2006
Heinrich, D., Hergt, M.: dtv-Atlas zur Ökologie. dtv, München, 3.A. 1994
Helmich, U. :Biomembranen. http://www.u-helmich.de/bio/cyt/reihe03/membran01.html , 2001/2005
Höffeler, F.: Bildatlas Cytologie. Harri Deutsch, Frankfurt a. M. 2003
Kattmann, U. (Hrsg.): Bioplanet Erde. UB 299 (Jg. 28), 2004
Krupinska, K., Desel, C., Mulisch: Stromuli – Plastidenbrücken im Netzwerk der Zelle. In: Biologie in unserer Zeit 40/3: S. 162–17, 2010,
Pott, R.: Allgemeine Geobotanik: Biogeosysteme und Biodiversität. Springer, Berlin 2005
Probst, W., Schuchardt, P. (Hrsg.): Biologie Ausgabe B. Duden-Paetec, Berlin/Frankfurt a.M. 2007
Probst, W. (Hrsg.): Miteinander- Beziehungennund Wechselwirkungen. UB 280 (Jg. 26), 2002
Probst, W. (Hrsg.): Ameisen und Termiten. UB 306 (Jg. 29), 2006
Rottmann, S.: Hier geht´s an die Nieren. UB 313 (Jg. 30), S.30-37, 2006
Ruppert, W. (Hrsg.): Struktur und Funktion. UB 232 (Jg.22), 1998
Simons, K., Ikonen, E. : Functional rafts in cell membranes. Nature 387, p. 569–572. 1997.
Suzuki, K. et al.: Rapid Hop Diffusion of a G-Protein-Coupled Receptor in the Plasma Membrane as Revealed by Single-Molecule Techniques. Biophysical Journal 88:3659-3680 (2005) http://www.biophysj.org/cgi/content/full/88/5/3659#FIG1
Wächtershäuser, G.: Evolution of the first metabolic cycles. Proceedings of the National Academy of Sciences, Vol. 87, Jan. 1990, p. 200–204
Wächtershäuser, G.: Origin of Life: Life as we don’t know it. Science 289 (5483), 25. August 2000, S. 1307–1308
Walter, H.: Die ökologischen Systeme der Kontinente (Biogeosphäre). G. Fischer, Stuttgart 1976
Walter, H.: Allgemeine Geobotanik. Ulmer, Stuttgart 1973
Whitfield, J.: Born in a watery commune. Nature 427, 19. Febr. 2004, p.674-676
Whittaker, R.H.: Communities an ecosystems. Macmillan, London/New York, 2.ed. 1975
Extrazelluläre Matrix:
http://www.unifr.ch/anatomy/elearningfree/allemand/bindegewebe/sfa/d-sfa.php
http://www.uni-tuebingen.de/uni/kxm/Courses/documents/GV0607ECM.pdf
Genzyme Deutschland: Einführung in das Krankheitsbild des Morbus Pompe. Neu-Isenburg, 2008. http://www.genzyme.de/thera/pompe/de_p_tp_thera-pompe.asp
Arealkunde
http://weinmannia.botanik.uni-hohenheim.de/Studienunterlagen_Dalitz/PDF/Arealkunde.pdf
Biotopbrücken
http://www.umweltbundesamt.at/umweltschutz/naturschutz/lebensraumschutz/vernetzung/lrv_empfehlungen/








